

Which Organelle Carries Out Photosynthesis
All animals and most microorganisms rely on the continual uptake of large amounts of organic compounds from their environment. These compounds are used to provide both the carbon skeletons for biosynthesis and the metabolic energy that drives cellular processes. Information technology is believed that the first organisms on the primitive Earth had access to an abundance of the organic compounds produced by geochemical processes, but that nearly of these original compounds were used up billions of years ago. Since that time, the vast bulk of the organic materials required by living cells have been produced by photosynthetic organisms, including many types of photosynthetic bacteria.
The most advanced photosynthetic bacteria are the cyanobacteria, which accept minimal nutrient requirements. They use electrons from h2o and the free energy of sunlight when they catechumen atmospheric CO2 into organic compounds—a procedure called carbon fixation. In the course of splitting water [in the overall reaction nH2O + northwardCO2

(CH2O) n + nO2], they also liberate into the atmosphere the oxygen required for oxidative phosphorylation. Every bit we see in this section, it is thought that the evolution of cyanobacteria from more primitive photosynthetic leaner somewhen fabricated possible the development of abundant aerobic life forms.
In plants and algae, which adult much later, photosynthesis occurs in a specialized intracellular organelle—the chloroplast. Chloroplasts perform photosynthesis during the daylight hours. The immediate products of photosynthesis, NADPH and ATP, are used by the photosynthetic cells to produce many organic molecules. In plants, the products include a low-molecular-weight sugar (commonly sucrose) that is exported to meet the metabolic needs of the many nonphotosynthetic cells of the organism.
Biochemical and genetic show strongly advise that chloroplasts are descendants of oxygen-producing photosynthetic leaner that were endocytosed and lived in symbiosis with primitive eucaryotic cells. Mitochondria are as well mostly believed to be descended from an endocytosed bacterium. The many differences between chloroplasts and mitochondria are thought to reflect their unlike bacterial ancestors, too equally their subsequent evolutionary divergence. Nevertheless, the key mechanisms involved in calorie-free-driven ATP synthesis in chloroplasts are very similar to those that we have already discussed for respiration-driven ATP synthesis in mitochondria.
The Chloroplast Is One Member of the Plastid Family unit of Organelles
Chloroplasts are the most prominent members of the plastid family of organelles. Plastids are nowadays in all living establish cells, each cell type having its own feature complement. All plastids share certain features. Most notably, all plastids in a particular establish species incorporate multiple copies of the same relatively pocket-size genome. In addition, each is enclosed past an envelope equanimous of ii concentric membranes.
As discussed in Chapter 12 (see Figure 12-iii), all plastids develop from proplastids, minor organelles in the immature cells of plant meristems (Effigy 14-33A). Proplastids develop according to the requirements of each differentiated prison cell, and the blazon that is present is determined in large function by the nuclear genome. If a leaf is grown in darkness, its proplastids enlarge and develop into etioplasts, which have a semicrystalline array of internal membranes containing a yellow chlorophyll precursor instead of chlorophyll. When exposed to light, the etioplasts quickly develop into chloroplasts by converting this precursor to chlorophyll and by synthesizing new membrane pigments, photosynthetic enzymes, and components of the electron-transport chain.

Figure 14-33
Plastid diversity. (A) A proplastid from a root tip prison cell of a edible bean plant. Annotation the double membrane; the inner membrane has also generated the relatively sparse internal membranes present. (B) Three amyloplasts (a grade of leucoplast), or starch-storing (more...)
Leucoplasts are plastids present in many epidermal and internal tissues that exercise not become green and photosynthetic. They are picayune more than enlarged proplastids. A common course of leucoplast is the amyloplast (Figure 14-33B), which accumulates the polysaccharide starch in storage tissues—a source of saccharide for future use. In some plants, such equally potatoes, the amyloplasts tin abound to be every bit big as an average fauna prison cell.
It is important to realize that plastids are non just sites for photosynthesis and the deposition of storage materials. Plants have likewise used their plastids to compartmentalize their intermediary metabolism. Purine and pyrimidine synthesis, most amino acid synthesis, and all of the fatty acrid synthesis of plants takes place in the plastids, whereas in brute cells these compounds are produced in the cytosol.
Chloroplasts Resemble Mitochondria But Have an Extra Compartment
Chloroplasts carry out their free energy interconversions past chemiosmotic mechanisms in much the same way that mitochondria do. Although much larger (Figure 14-34A), they are organized on the same principles. They accept a highly permeable outer membrane; a much less permeable inner membrane, in which membrane transport proteins are embedded; and a narrow intermembrane space in betwixt. Together, these membranes form the chloroplast envelope (Figure 14-34B,C). The inner membrane surrounds a big infinite called the stroma, which is analogous to the mitochondrial matrix and contains many metabolic enzymes. Similar the mitochondrion, the chloroplast has its ain genome and genetic system. The stroma therefore also contains a special set of ribosomes, RNAs, and the chloroplast DNA.
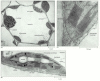
Figure fourteen-34
Electron micrographs of chloroplasts. (A) In a wheat leaf cell, a thin rim of cytoplasm—containing chloroplasts, the nucleus, and mitochondria—surrounds a large vacuole. (B) A sparse department of a single chloroplast, showing the chloroplast (more...)
There is, however, an of import difference between the organisation of mitochondria and that of chloroplasts. The inner membrane of the chloroplast is not folded into cristae and does not contain electron-send chains. Instead, the electron-transport bondage, photosynthetic low-cal-capturing systems, and ATP synthase are all contained in the thylakoid membrane, a tertiary distinct membrane that forms a set of flattened disclike sacs, the thylakoids (Effigy xiv-35). The lumen of each thylakoid is thought to be continued with the lumen of other thylakoids, thereby defining a tertiary internal compartment called the thylakoid space, which is separated past the thylakoid membrane from the stroma that surrounds it.

Figure 14-35
The chloroplast. This photosynthetic organelle contains three singled-out membranes (the outer membrane, the inner membrane, and the thylakoid membrane) that ascertain three separate internal compartments (the intermembrane space, the stroma, and the thylakoid (more...)
The structural similarities and differences between mitochondria and chloroplasts are illustrated in Figure fourteen-36. The head of the chloroplast ATP synthase, where ATP is made, protrudes from the thylakoid membrane into the stroma, whereas information technology protrudes into the matrix from the inner mitochondrial membrane.

Effigy 14-36
A mitochondrion and chloroplast compared. A chloroplast is generally much larger than a mitochondrion and contains, in addition to an outer and inner membrane, a thylakoid membrane enclosing a thylakoid space. Unlike the chloroplast inner membrane, the (more than...)
Chloroplasts Capture Energy from Sunlight and Apply Information technology to Fix Carbon
The many reactions that occur during photosynthesis in plants can be grouped into 2 broad categories:
- one.
-
In the photosynthetic electron-transfer reactions (also called the "light reactions"), energy derived from sunlight energizes an electron in the greenish organic paint chlorophyll, enabling the electron to move forth an electron-transport chain in the thylakoid membrane in much the aforementioned style that an electron moves along the respiratory concatenation in mitochondria. The chlorophyll obtains its electrons from water (H2O), producing O2 equally a by-production. During the electron-transport process, H+ is pumped across the thylakoid membrane, and the resulting electrochemical proton gradient drives the synthesis of ATP in the stroma. Equally the final step in this series of reactions, high-free energy electrons are loaded (together with H+) onto NADP+, converting it to NADPH. All of these reactions are confined to the chloroplast.
- 2.
-
In the carbon-fixation reactions (too chosen the "night reactions"), the ATP and the NADPH produced by the photosynthetic electron-transfer reactions serve equally the source of energy and reducing ability, respectively, to drive the conversion of CO2 to sugar. The carbon-fixation reactions, which begin in the chloroplast stroma and continue in the cytosol, produce sucrose and many other organic molecules in the leaves of the plant. The sucrose is exported to other tissues as a source of both organic molecules and energy for growth.
Thus, the germination of ATP, NADPH, and O2 (which requires calorie-free free energy directly) and the conversion of COtwo to carbohydrate (which requires light free energy but indirectly) are separate processes (Figure xiv-37), although elaborate feedback mechanisms interconnect the 2. Several of the chloroplast enzymes required for carbon fixation, for example, are inactivated in the dark and reactivated by low-cal-stimulated electron-transport processes.
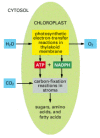
Figure xiv-37
The reactions of photosynthesis in a chloroplast. Water is oxidized and oxygen is released in the photosynthetic electron-transfer reactions, while carbon dioxide is assimilated (fixed) to produce sugars and a diverseness of other organic molecules in the (more...)
Carbon Fixation Is Catalyzed by Ribulose Bisphosphate Carboxylase
We take seen earlier in this chapter how cells produce ATP by using the large amount of gratis free energy released when carbohydrates are oxidized to CO2 and H2O. Clearly, therefore, the reverse reaction, in which CO2 and H2O combine to make saccharide, must exist a very unfavorable 1 that can but occur if it is coupled to other, very favorable reactions that drive it.
The fundamental reaction of carbon fixation, in which an atom of inorganic carbon is converted to organic carbon, is illustrated in Figure 14-38: COtwo from the atmosphere combines with the 5-carbon compound ribulose 1,5-bisphosphate plus h2o to yield two molecules of the three-carbon compound 3-phosphoglycerate. This "carbon-fixing" reaction, which was discovered in 1948, is catalyzed in the chloroplast stroma past a large enzyme called ribulose bisphosphate carboxylase. Since each molecule of the complex works sluggishly (processing only virtually 3 molecules of substrate per second compared to 1000 molecules per second for a typical enzyme), many enzyme molecules are needed. Ribulose bisphosphate carboxylase often constitutes more fifty% of the full chloroplast protein, and it is idea to exist the about abundant protein on Globe.
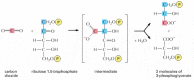
Figure 14-38
The initial reaction in carbon fixation. This reaction, in which carbon dioxide is converted into organic carbon, is catalyzed in the chloroplast stroma past the abundant enzyme ribulose bisphosphate carboxylase. The production is three-phosphoglycerate, which (more than...)
Three Molecules of ATP and Two Molecules of NADPH Are Consumed for Each CO2 Molecule That Is Fixed
The bodily reaction in which CO2 is stock-still is energetically favorable because of the reactivity of the energy-rich compound ribulose 1,5-bisphosphate, to which each molecule of CO2 is added (run into Figure 14-38). The elaborate metabolic pathway that produces ribulose one,5-bisphosphate requires both NADPH and ATP; it was worked out in one of the first successful applications of radioisotopes as tracers in biochemistry. This carbon-fixation wheel (also called the Calvin cycle) is outlined in Figure xiv-39. It starts when 3 molecules of CO2 are stock-still by ribulose bisphosphate carboxylase to produce half dozen molecules of 3-phosphoglycerate (containing 6 × 3 = xviii carbon atoms in all: 3 from the COtwo and fifteen from ribulose 1,v-bisphosphate). The 18 carbon atoms and so undergo a bicycle of reactions that regenerates the iii molecules of ribulose 1,5-bisphosphate used in the initial carbon-fixation step (containing iii × 5 = fifteen carbon atoms). This leaves one molecule of glyceraldehyde 3-phosphate (3 carbon atoms) every bit the internet gain.
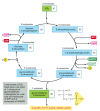
Figure 14-39
The carbon-fixation cycle, which forms organic molecules from COii and H2O. The number of carbon atoms in each type of molecule is indicated in the white box. There are many intermediates between glyceraldehyde iii-phosphate and ribulose 5-phosphate, only (more than...)
A total of 3 molecules of ATP and ii molecules of NADPH are consumed for each CO2 molecule converted into carbohydrate. The net equation is:

Thus, both phosphate-bail energy (every bit ATP) and reducing power (every bit NADPH) are required for the germination of organic molecules from CO2 and H2O. Nosotros render to this important point subsequently.
The glyceraldehyde three-phosphate produced in chloroplasts by the carbon-fixation wheel is a three-carbon sugar that also serves as a central intermediate in glycolysis. Much of information technology is exported to the cytosol, where information technology tin be converted into fructose 6-phosphate and glucose one-phosphate by the reversal of several reactions in glycolysis (see Panel 2-8, pp. 124–125). The glucose one-phosphate is then converted to the carbohydrate nucleotide UDP-glucose, and this combines with the fructose 6-phosphate to grade sucrose phosphate, the immediate forerunner of the disaccharide sucrose. Sucrose is the major grade in which sugar is transported between plant cells: just equally glucose is transported in the blood of animals, sucrose is exported from the leaves via vascular bundles, providing the saccharide required past the rest of the plant.
Virtually of the glyceraldehyde 3-phosphate that remains in the chloroplast is converted to starch in the stroma. Like glycogen in animal cells, starch is a large polymer of glucose that serves as a carbohydrate reserve (see Figure 14-33B). The production of starch is regulated so that information technology is produced and stored as large grains in the chloroplast stroma during periods of excess photosynthetic capacity. This occurs through reactions in the stroma that are the reverse of those in glycolysis: they convert glyceraldehyde three-phosphate to glucose i-phosphate, which is so used to produce the saccharide nucleotide ADP-glucose, the firsthand precursor of starch. At dark the starch is broken down to help support the metabolic needs of the plant. Starch provides an important part of the diet of all animals that eat plants.
Carbon Fixation in Some Plants Is Compartmentalized to Facilitate Growth at Low CO2 Concentrations
Although ribulose bisphosphate carboxylase preferentially adds COtwo to ribulose i,5-bisphosphate, it can apply O2 as a substrate in place of CO2, and if the concentration of CO2 is depression, information technology volition add Otwo to ribulose 1,five-bisphosphate instead (see Figure 14-38). This is the get-go stride in a pathway called photorespiration, whose ultimate effect is to use upwards O2 and liberate COtwo without the production of useful energy stores. In many plants, about one-tertiary of the COii fixed is lost again as COtwo because of photorespiration.
Photorespiration can exist a serious liability for plants in hot, dry weather, which crusade them to shut their stomata (the gas commutation pores in their leaves) to avoid excessive water loss. This in plow causes the CO2 levels in the leaf to fall precipitously, thereby favoring photorespiration. A special adaptation, however, occurs in the leaves of many plants, such as corn and carbohydrate pikestaff that alive in hot, dry environments. In these plants, the carbon-fixation cycle occurs just in the chloroplasts of specialized bundle-sheath cells, which incorporate all of the plant's ribulose bisphosphate carboxylase. These cells are protected from the air and are surrounded past a specialized layer of mesophyll cells that use the free energy harvested by their chloroplasts to "pump" COtwo into the parcel-sheath cells. This supplies the ribulose bisphosphate carboxylase with a high concentration of COtwo, thereby greatly reducing photorespiration.
The COii pump is produced past a reaction cycle that begins in the cytosol of the mesophyll cells. A CO2-fixation step is catalyzed past an enzyme that binds carbon dioxide (as bicarbonate) and combines information technology with an activated three-carbon molecule to produce a four-carbon molecule. The four-carbon molecule diffuses into the bundle-sheath cells, where information technology is broken down to release the CO2 and generate a molecule with three carbons. The pumping cycle is completed when this three-carbon molecule is returned to the mesophyll cells and converted back to its original activated form. Because the CO2 is initially captured by converting it into a compound containing four carbons, the CO2-pumping plants are called Civ plants. All other plants are called C3 plants because they capture CO2 into the iii-carbon chemical compound 3-phosphoglycerate (Figure 14-40).

Figure 14-40
Comparative leafage anatomy in a Ciii plant and a Civ establish. The cells with green cytosol in the leafage interior comprise chloroplasts that perform the normal carbon-fixation cycle. In C4 plants, the mesophyll cells are specialized for CO2 pumping rather than (more than...)
As for whatsoever vectorial send process, pumping CO2 into the bundle-sheath cells in C4 plants costs free energy. In hot, dry out environments, notwithstanding, this cost tin can exist much less than the free energy lost by photorespiration in Ciii plants, and so C4 plants accept a potential advantage. Moreover, considering Cfour plants can perform photosynthesis at a lower concentration of CO2 within the foliage, they need to open their stomata less often and therefore can fix well-nigh twice every bit much net carbon equally C3 plants per unit of h2o lost. Although the vast majority of plant species are C3 plants, C4 plants such as corn and sugar pikestaff are much more than effective at converting sunlight energy into biomass than C3 plants such as cereal grains. They are therefore of special importance in earth agriculture.
Photosynthesis Depends on the Photochemistry of Chlorophyll Molecules
Having discussed the carbon-fixation reactions, we now return to the question of how the photosynthetic electron-transfer reactions in the chloroplast generate the ATP and the NADPH needed to drive the production of carbohydrates from COii and H2O. The required free energy is derived from sunlight absorbed by chlorophyll molecules (Effigy 14-41). The process of energy conversion begins when a chlorophyll molecule is excited by a quantum of light (a photon) and an electron is moved from one molecular orbital to some other of higher free energy. As illustrated in Figure 14-42, such an excited molecule is unstable and tends to return to its original, unexcited state in one of three ways:
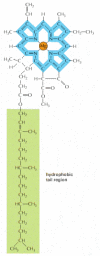
Effigy 14-41
The structure of chlorophyll. A magnesium atom is held in a porphyrin ring, which is related to the porphyrin ring that binds iron in heme (run into Figure 14-22). Electrons are delocalized over the bonds shown in blue.

Figure 14-42
Three ways for an excited chlorophyll molecule to return to its original, unexcited state. The light energy captivated by an isolated chlorophyll molecule is completely released as calorie-free and heat by procedure one. In photosynthesis, past dissimilarity, chlorophylls (more than...)
- 1.
-
Past converting the extra free energy into heat (molecular motions) or to some combination of heat and calorie-free of a longer wavelength (fluorescence), which is what happens when light energy is absorbed by an isolated chlorophyll molecule in solution.
- two.
-
By transferring the energy—but not the electron—directly to a neighboring chlorophyll molecule past a process called resonance free energy transfer.
- iii.
-
By transferring the high-energy electron to another nearby molecule, an electron acceptor, and so returning to its original land by taking up a low-energy electron from some other molecule, an electron donor.
The last two mechanisms are exploited in the procedure of photosynthesis.
A Photosystem Consists of a Reaction Middle Plus an Antenna Complex
Multiprotein complexes called photosystems catalyze the conversion of the lite free energy captured in excited chlorophyll molecules to useful forms. A photosystem consists of two closely linked components: an antenna complex, consisting of a big set of pigment molecules that capture light energy and feed information technology to the reaction center; and a photochemical reaction eye, consisting of a complex of proteins and chlorophyll molecules that enable lite energy to be converted into chemical energy (Effigy 14-43).
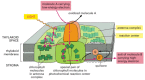
Figure 14-43
The antenna complex and photochemical reaction center in a photosystem. The antenna circuitous is a collector of light energy in the form of excited electrons. The free energy of the excited electrons is funneled, through a series of resonance free energy transfers, (more...)
The antenna complex is important for capturing low-cal. In chloroplasts it consists of a number of distinct membrane poly peptide complexes (known as light-harvesting complexes); together, these proteins bind several hundred chlorophyll molecules per reaction center, orienting them precisely in the thylakoid membrane. Depending on the plant, different amounts of accessory pigments called carotenoids, which protect the chlorophylls from oxidation and can help collect light of other wavelengths, are too located in each circuitous. When a chlorophyll molecule in the antenna complex is excited, the free energy is rapidly transferred from one molecule to some other by resonance energy transfer until it reaches a special pair of chlorophyll molecules in the photochemical reaction center. Each antenna complex thereby acts equally a funnel, collecting light energy and directing information technology to a specific site where it tin exist used effectively (see Effigy 14-43).
The photochemical reaction centre is a transmembrane protein-pigment complex that lies at the heart of photosynthesis. It is thought to have evolved more than iii billion years ago in primitive photosynthetic leaner. The special pair of chlorophyll molecules in the reaction center acts as an irreversible trap for excitation quanta because its excited electron is immediately passed to a chain of electron acceptors that are precisely positioned every bit neighbors in the same protein complex (Figure 14-44). By moving the high-energy electron rapidly away from the chlorophylls, the photochemical reaction center transfers it to an environment where it is much more stable. The electron is thereby suitably positioned for subsequent reactions, which require more time to consummate.

Figure 14-44
The arrangement of the electron carriers in a bacterial photochemical reaction center, equally determined by x-ray crystallography. The pigment molecules shown are held in the interior of a transmembrane protein and are surrounded past the lipid bilayer of the (more than...)
In a Reaction Center, Light Energy Captured by Chlorophyll Creates a Strong Electron Donor from a Weak One
The electron transfers involved in the photochemical reactions just outlined accept been analyzed extensively by rapid spectroscopic methods. An enormous corporeality of detailed information is available for the photosystem of majestic bacteria, which is somewhat simpler than the evolutionarily related photosystems in chloroplasts. The reaction eye in this photosystem is a large protein-pigment complex that can be solubilized with detergent and purified in active form. In 1985, its complete three-dimensional structure was determined by ten-ray crystallography (see Figure 10-38). This structure, combined with kinetic data, provides the best motion picture we have of the initial electron-transfer reactions that underlie photosynthesis.
The sequence of electron transfers that take place in the reaction center of regal bacteria is shown in Effigy xiv-45. As outlined previously for the general case (see Figure 14-43), low-cal causes a net electron transfer from a weak electron donor (a molecule with a strong affinity for electrons) to a molecule that is a strong electron donor in its reduced course. The excitation energy in chlorophyll that would usually be released equally fluorescence or rut is thereby used instead to create a strong electron donor (a molecule carrying a high-energy electron) where none had been before. In the purple bacterium, the weak electron donor used to fill the electron-deficient hole created past a light-induced charge separation is a cytochrome (see orange box in Figure 14-45); the stiff electron donor produced is a quinone. In the chloroplasts of higher plants, a quinone is similarly produced. Nonetheless, as nosotros discuss next, water serves every bit the initial weak electron donor, which is why oxygen gas is released by photosynthesis in plants.

Figure 14-45
The electron transfers that occur in the photochemical reaction center of a purple bacterium. A like fix of reactions occurs in the evolutionarily related photosystem II in plants. At the superlative left is an orientating diagram showing the molecules that (more than...)
Noncyclic Photophosphorylation Produces Both NADPH and ATP
Photosynthesis in plants and cyanobacteria produces both ATP and NADPH directly by a two-step process chosen noncyclic photophosphorylation. Because two photosystems—chosen photosystems I and 2—are used in series to energize an electron, the electron can be transferred all the way from water to NADPH. As the high-energy electrons pass through the coupled photosystems to generate NADPH, some of their free energy is siphoned off for ATP synthesis.
The first of the ii photosystems—paradoxically called photosystem II for historical reasons—has the unique ability to withdraw electrons from water. The oxygens of two water molecules bind to a cluster of manganese atoms in a poorly understood water-splitting enzyme. This enzyme enables electrons to be removed i at a time from the water, every bit required to fill up the electron-deficient holes created by light in chlorophyll molecules in the reaction heart. As presently as four electrons have been removed from the two water molecules (requiring four quanta of light), Oii is released. Photosystem 2 thus catalyzes the reaction 2HtwoO + 4 photons → 4H+ + 4eastward - + O2. As we discussed for the electron-send concatenation in mitochondria, which uses Otwo and produces water, the mechanism ensures that no partly oxidized water molecules are released as dangerous, highly reactive oxygen radicals. Essentially all the oxygen in the Earth'south temper has been produced in this way.
The core of the reaction centre in photosystem Ii is homologous to the bacterial reaction center just described, and information technology likewise produces strong electron donors in the class of reduced quinone molecules dissolved in the lipid bilayer of the membrane. The quinones pass their electrons to a H+ pump chosen the cytochrome b6-f complex, which resembles the cytochrome b-c 1 complex in the respiratory concatenation of mitochondria. The cytochrome b half dozen -f complex pumps H+ into the thylakoid space across the thylakoid membrane (or out of the cytosol across the plasma membrane in cyanobacteria), and the resulting electrochemical gradient drives the synthesis of ATP past an ATP synthase (Effigy 14-46).

Figure 14-46
Electron flow during photosynthesis in the thylakoid membrane. The mobile electron carriers in the concatenation are plastoquinone (which closely resembles the ubiquinone of mitochondria), plastocyanin (a small copper-containing protein), and ferredoxin (a small (more...)
The final electron acceptor in this electron-ship chain is the 2d photosystem, photosystem I, which accepts an electron into the electron-deficient hole created past light in the chlorophyll molecule in its reaction center. Each electron that enters photosystem I is finally additional to a very loftier-free energy level that allows it to exist passed to the iron-sulfur center in ferredoxin and then to NADP+ to generate NADPH (Effigy fourteen-47).

Effigy xiv-47
Changes in redox potential during photosynthesis. The redox potential for each molecule is indicated by its position along the vertical centrality. In photosystem Two, the excited reaction centre chlorophyll has a redox potential high enough to withdraw electrons (more than...)
The scheme for photosynthesis merely discussed is known equally the Z scheme. By means of its ii electron-energizing steps, ane catalyzed past each photosystem, an electron is passed from h2o, which ordinarily holds on to its electrons very tightly (redox potential = +820 mV), to NADPH, which normally holds on to its electrons loosely (redox potential = -320 mV). There is non enough free energy in a single breakthrough of visible light to energize an electron all the fashion from the bottom of photosystem Two to the top of photosystem I, which is presumably the free energy change required to pass an electron efficiently from water to NADP+. The utilize of two separate photosystems in series ways that the energy from ii quanta of low-cal is available for this purpose. In addition, there is enough energy left over to enable the electron-send chain that links the two photosystems to pump H+ across the thylakoid membrane (or the plasma membrane of cyanobacteria), so that the ATP synthase can harness some of the light-derived energy for ATP product.
Chloroplasts Can Brand ATP by Cyclic Photophosphorylation Without Making NADPH
In the noncyclic photophosphorylation scheme just discussed, loftier-free energy electrons leaving photosystem II are harnessed to generate ATP and are passed on to photosystem I to drive the production of NADPH. This produces slightly more than than 1 molecule of ATP for every pair of electrons that passes from HtwoO to NADP+ to generate a molecule of NADPH. But ane.5 molecules of ATP per NADPH are needed for carbon fixation (run across Figure fourteen-39). To produce extra ATP, the chloroplasts in some species of plants can switch photosystem I into a cyclic mode so that information technology produces ATP instead of NADPH. In this procedure, called circadian photophosphorylation, the high-energy electrons from photosystem I are transferred to the cytochrome b 6 -f complex rather than being passed on to NADP+. From the b 6 -f circuitous, the electrons are passed dorsum to photosystem I at a low energy. The only internet result, also the conversion of some light energy to heat, is that H+ is pumped across the thylakoid membrane past the b vi -f complex every bit electrons laissez passer through it, thereby increasing the electrochemical proton gradient that drives the ATP synthase. (This is analogous to the correct side of the diagram for purple nonsulfur bacteria in Effigy 14-71, below.)
To summarize, circadian photophosphorylation involves merely photosystem I, and it produces ATP without the formation of either NADPH or O2. The relative activities of circadian and noncyclic electron flows can be regulated by the prison cell to determine how much light energy is converted into reducing ability (NADPH) and how much into loftier-energy phosphate bonds (ATP).
Photosystems I and 2 Take Related Structures, and Also Resemble Bacterial Photosystems
The mechanisms of fundamental prison cell processes such equally DNA replication or respiration more often than not turn out to be the same in eucaryotic cells and in bacteria, even though the number of poly peptide components involved is considerably greater in eucaryotes. Eucaryotes evolved from procaryotes, and the additional proteins presumably were selected for during evolution because they provided an extra degree of efficiency and/or regulation that was useful to the prison cell.
Photosystems provide a clear case of this blazon of evolution. Photosystem II, for example, is formed from more than 25 dissimilar protein subunits, creating a large assembly in the thylakoid membrane with a mass of about 1 million daltons. The atomic structures of the eucaryotic photosystems are existence revealed by a combination of electron and x-ray crystallography. The job is difficult considering the complexes are large and embedded in the lipid bilayer. Nevertheless, as illustrated in Effigy 14-48, the close relationship of photosystem I, photosystem II, and the photochemical reaction center of purple bacteria has been clearly demonstrated from these atomic-level analyses.

Figure fourteen-48
Three types of photosynthetic reaction centers compared. Pigments involved in lite harvesting are colored light-green; those involved in the central photochemical events are colored red. (A) The photochemical reaction centre of purple bacteria, whose detailed (more than...)
The Proton-Motive Strength Is the Aforementioned in Mitochondria and Chloroplasts
The presence of the thylakoid space separates a chloroplast into three rather than the two internal compartments of a mitochondrion. The net effect of H+ translocation in the two organelles is, all the same, like. Every bit illustrated in Figure 14-49, in chloroplasts, H+ is pumped out of the stroma (pH 8) into the thylakoid space (pH ~5), creating a gradient of three–3.five pH units. This represents a proton-motive forcefulness of almost 200 mV across the thylakoid membrane, and it drives ATP synthesis past the ATP synthase embedded in this membrane. The strength is the aforementioned as that across the inner mitochondrial membrane, merely nearly all of it is contributed by the pH gradient rather than by a membrane potential, unlike the case in mitochondria.

Figure 14-49
A comparison of the period of H+ and the orientation of the ATP synthase in mitochondria and chloroplasts. Those compartments with like pH values have been colored the same. The proton-motive force across the thylakoid membrane consists near entirely (more...)
Similar the stroma, the mitochondrial matrix has a pH of about viii. This is created by pumping H+ out of the mitochondrion into the cytosol (pH ~7) rather than into an interior infinite in the organelle. Thus, the pH slope is relatively small, and well-nigh of the proton-motive force across the inner mitochondrial membrane is instead caused by the resulting membrane potential (see Effigy 14-13).
For both mitochondria and chloroplasts, the catalytic site of the ATP synthase is at a pH of about 8 and is located in a big organelle compartment (matrix or stroma) that is packed full of soluble enzymes. Consequently, information technology is here that all of the organelle's ATP is made (run across Figure 14-49).
Carrier Proteins in the Chloroplast Inner Membrane Control Metabolite Exchange with the Cytosol
If chloroplasts are isolated in a way that leaves their inner membrane intact, this membrane tin can be shown to have a selective permeability, reflecting the presence of specific carrier proteins. Most notably, much of the glyceraldehyde 3-phosphate produced by CO2 fixation in the chloroplast stroma is transported out of the chloroplast by an efficient antiport arrangement that exchanges three-carbon sugar phosphates for an inward flux of inorganic phosphate.
Glyceraldehyde 3-phosphate ordinarily provides the cytosol with an arable source of saccharide, which is used past the jail cell as the starting betoken for many other biosyntheses—including the product of sucrose for export. But this is non all that this molecule provides. In one case the glyceraldehyde 3-phosphate reaches the cytosol, it is readily converted (by part of the glycolytic pathway) to 1,three-phosphoglycerate and then iii-phosphoglycerate (see p. 97), generating one molecule of ATP and one of NADH. (A similar ii-pace reaction, but working in reverse, forms glyceraldehyde 3-phosphate in the carbon-fixation bike; come across Figure 14-39.) As a outcome, the export of glyceraldehyde iii-phosphate from the chloroplast provides non simply the chief source of stock-still carbon to the remainder of the prison cell, just also the reducing power and ATP needed for metabolism outside the chloroplast.
Chloroplasts Besides Perform Other Crucial Biosyntheses
The chloroplast performs many biosyntheses in improver to photosynthesis. All of the cell'south fatty acids and a number of amino acids, for example, are made by enzymes in the chloroplast stroma. Similarly, the reducing power of light-activated electrons drives the reduction of nitrite (NO2 -) to ammonia (NH3) in the chloroplast; this ammonia provides the institute with nitrogen for the synthesis of amino acids and nucleotides. The metabolic importance of the chloroplast for plants and algae therefore extends far beyond its role in photosynthesis.
Summary
Chloroplasts and photosynthetic bacteria obtain high-energy electrons by means of photosystems that capture the electrons that are excited when sunlight is absorbed by chlorophyll molecules. Photosystems are composed of an antenna complex that funnels free energy to a photochemical reaction center, where a precisely ordered complex of proteins and pigments allows the energy of an excited electron in chlorophyll to be captured by electron carriers. The all-time-understood photochemical reaction center is that of purple photosynthetic bacteria, which contain only a single photosystem. In dissimilarity, there are two singled-out photosystems in chloroplasts and blue-green alga. The two photosystems are normally linked in serial, and they transfer electrons from water to NADP+ to form NADPH, with the concomitant product of a transmembrane electrochemical proton slope. In these linked photosystems, molecular oxygen (O2) is generated every bit a past-product of removing four low-free energy electrons from 2 specifically positioned water molecules.
Compared with mitochondria, chloroplasts have an boosted internal membrane (the thylakoid membrane) and a tertiary internal space (the thylakoid space). All electron-transport processes occur in the thylakoid membrane: to make ATP, H+ is pumped into the thylakoid infinite, and a backflow of H+ through an ATP synthase then produces the ATP in the chloroplast stroma. This ATP is used in conjunction with the NADPH made past photosynthesis to bulldoze a large number of biosynthetic reactions in the chloroplast stroma, including the all-of import carbon-fixation cycle, which creates saccharide from CO2. Along with another of import chloroplast products, this carbohydrate is exported to the cell cytosol, where—equally glyceraldehyde 3-phosphate—it provides organic carbon, ATP, and reducing power to the remainder of the cell.

Which Organelle Carries Out Photosynthesis,
Source: https://www.ncbi.nlm.nih.gov/books/NBK26819/
Posted by: coatestherds.blogspot.com
0 Response to "Which Organelle Carries Out Photosynthesis"
Post a Comment